
FACULTAD DE CIENCIAS
DEPARTAMENTO DE BIOLOGÍA
LABORATORIO DE BIOLOGIA DE PROTOZOARIOS
(L.B.P)
PROTOZOOLOGIA
PROTOZOOLOGIA
volver a indproto+ volver a proginfLBP
Generalidades y Sistematica: Los flagelados incluidos en
este género se caracterizan por: a) Son digenéticos
(dos
huesped, uno vertebrado y otro invertebrado) igual que el género
Trypanosoma, del cual se diferencian, entre otras características,
porque en su ciclo de vida sólo desarrollan
las formas amastigotes y las promastigotes. Desde el
punto de vista morfológico, el género que nos
ocupa, no se ha podido diferenciar del género leptomonas.
La separación se hace porque el último es
monogenético (un solo huesped, el
invertebrado). La forma del flagelado en
el vertebrado es de amastigote (cuerpo oval sin flagelo libre).
En el huésped invertebrado el cuerpo se alarga y desarrolla el
flagelo libre que sale por la parte anterior, pudiendo coexistir
otras formas intermedias asi como el amastigote. En la naturaleza
se encuentran infectados con este parásito los lagartos y mamíferos;
en estos últimos encontramos frecuentemente
infectados los cánidos, edentados, marsupiales, primates,
etc.
CICLO DE VIDA Y MORFOLOGIA:
El ciclo de vida de la leishmaniasis se realiza en dos huésped por alternancia entre los vectores flebotominos (en el viejo mundo) o Lutzomyia (en el nuevo mundo) y el vertebrado. Profundos cambios biológicos acompañan este ciclo, transformandose en amastigote dentro de los macrófagos de vertebrados y en un flagelado mótil, extracelular que habita el intestino de un insecto. Leishmania donovani (visceral) está adaptada a la transmisión hombre-vector-hombre (antroponosis). Mientras que otras especies se desrrollan en una zoonosis en los reservorios vertebrados, por ejemplo: roedores-vector-roedores; en estos casos la transmisión al hombre es relativamente fortuita y depende mayormente de contactos entre hombre y vector previamente infestados, por alimentarse estos sobre un huésped reservorio infectado. Diferentes leishmanias pueden tener diferentes reservorios específicos. Son parásitos del sistemá retículo endotelial del huésped vertebrado.
Morfología:
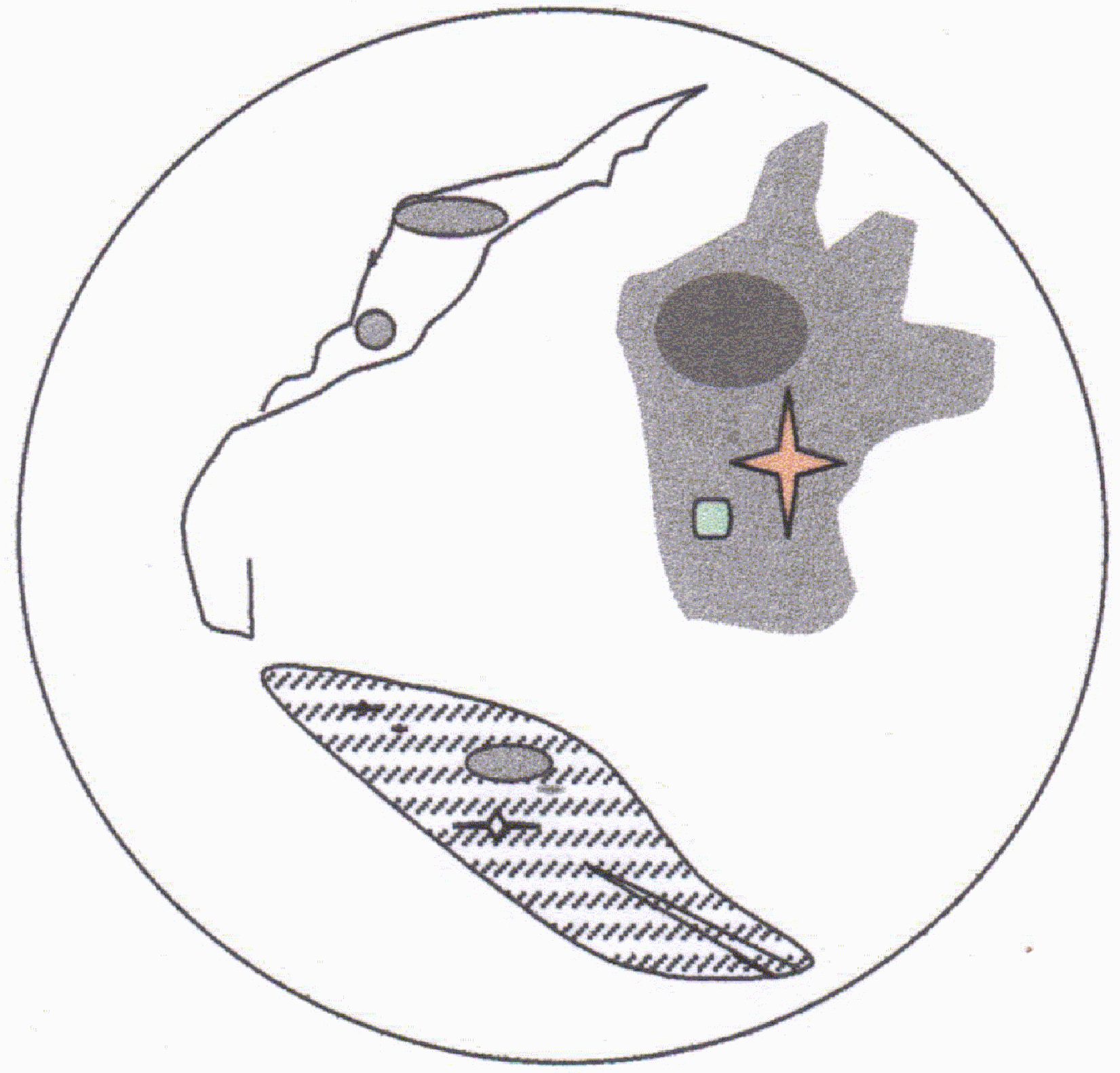
Microscopio de luz visible: a) el amastigote tiene cuerpo ovoide,
sin movimiento, mide entre 2 a 5 µm de longitud.
Contiene un núcleo central que se
tiñe de azul púrpura con tinsión
de Romanowsky, igual que el kinetoplasto. El axonema (rizoplasto) se puede
observar en algunas ocasiones. En la
naturaleza los amatigotes se encuentran intracelularmente
en los macrófagos (fagocitos mononucleares), cuando
por alguna causa se rompen los macrófagos entonces se observan
los amastigotes libres.
Se ha establecido que los amastigotes
de las diferentes leishmanias son morfologicamente
indistinguibles, al menos al microscopio de luz visible.
Esta generalización parece no ser del todo válida,
ya que algunos autores han descrito diferencias entre las leishmanias,
ya sea por el tamaño o por otras causas, por ejemplo:
la Leishmania mexicana presenta vacuolización,
Scorza & col. 1979, describieron una estructura birrefringente
(vacuolización), y que según ellos es característica
exclusiva de la leishmania de los andes venezolanos, Tovar y sus alrededores.
Se ha descrito la transformación morfológica de amastigotes a promastigotes in-vitro a 26°C.. El proceso reverso ocurre al entrar el flagelado en un macrófago a 37°C o 25°C.
El promastigote posee un solo flagelo, la longitud de su cuerpo es de 20µm y la de los flagelos es de igual longitud. Durante la división el kinetoplasto se divide en fase con el núcleo.
Microscopía electrónica: Diferentes autores han descrito tanto en amastigotes como en promastigotes, el núcleo con un nucleolo, mitocondrias con sus crestas, aparato de golgi, retículo endoplásmico y ribosomas como en otras células eucariotas. él sistema cinético flagelo, kinetoplasto y cuerpo basal se describió en las generalidades de trypanosomatidaes. No se han diferenciados, en la ultraestructura, a las diferentes leishmanias.
Taxonomía: Toda nueva técnica,
descubierta o aplicada en taxonomía,
produce una fuerte tentación para exaltarla, en
el primer momento, con todo entusiasmo; dándole la mayor importancia
por ser capaz de suministrar caracteres de mayor peso que
los hasta ahora usados. Esta tentación debe ser firmemente
resistida (Hawkes and Tucker, 1968).
Los primeros criterios usados para clasificar las
leishmanias se basaron mayormente en el aspecto clínico de las
leishmaniasis humana, siendo limitados sólo para éstas.
Históricamente se han dividido en las del Hemisferio del este
y las del hemisferio del oeste.
A. Leishmanias del Este:
Parece ser que el primero en observar parásitos de este género fue Cunningham cuando examinaba secciones histológicas del botón de oriente, en la India en 1885, quien lo confundió con un hongo (Slime fongi) y fue Barovsky quien estableció la naturaleza protozoológica del parásito cuando hizo su descripción del botón de oriente en Turkestán en 1898.
Leishman, descubrió el parásito responsable de la leishmaniasis visceral (Kala azar) en el hombre, en la India en 1903 y correctamente lo correlacionó con los tripanosomas. Laveran y Mesnil la denominaron Piroplasma donovani, hasta que Ross en 1903 le cambió el nombre por el de Leishmania donovani.
En 1903 Wright se refirió al agente causal del botón de oriente como Helcosoma tropicum, Mientras que Luhe fué el primero en asignar el nombre de Leismania trópica en 1906, pero pensaba que era un piroplasma. Rogers en 1904 y Nicolle en 1908 finalmente establecieron la naturaleza de Trypanosomatidae de L. donovani y L. tropica, respectivamente, reproduciendo el estado flagelar (promastigote) en cultivos in-vitro.
En el hemisferio oriental se han definido tres Leishmanias que infectan al hombre:
L. infantun, Nicolle 1908 - para la causante de viceralización en niños o "Kala azar" infantil en ciudades del Mediterráneo, así como en otras partes de Europa, Asia y Africa.
L. tropica (mayor), Yakimaff y Schokov en 1915, como la causa (agente etiológico) de la zoonosis de leishmaniasis cutánea en el Turkmen, república de la U.R.S.S.
L. aethiopica Bray et. al. (1973) para la leishmania causante de la leishmaniasis cutánea y la cutaneodifusa, en Etiopía y Kenya.
Hay la tendencia a considerar todas las leishmaniasis viscerales
como causadas por una sola especie, pero esto no tiene porque ser así,
ya que la misma clínica puede ser producida por diferentes
parásitos. Diferencias biológicas no han sido acumuladas,
en el futuro cuando se puedan determinar, entonces también
se podrán separar parásitos diferentes.
Leishmaniasis del Hemiferio Occidental:
Leishmaniasis cutánea:
En Sur América fue Lindenberg quien descubrió la leishmaniasis cutánea en Brasil (1909), encontró que se extendía desde Centro hasta Sur América, la única excepción parece ser chile.
Los primeros investigadores pensaron que se trataba de un solo parásito L. brasiliensis, Vianna 1912, pero luego se han podido encontrar algunas diferencias; se ha descrito L. peruviana, Velez (1913) el agente etiológico de la "UTA" en los Andes de Perú. La aparición, también, de L. enriettii en acures de laboratorio. Estos casos y otros, nos dicen de una multiplicidad de leishmanias en el Hemiferio Occidental.
Biagi (1953) y Floch (1954) refirieron a L. tropica mexicana como la causante de la úlcera del chiclero en Yucatán, Guatemala y Belize (Honduras ), a L. tropica braziliensis como la causante de leishmaniasis mucocutánea en Brasil, y la L. tropica guayanensis como causante de "pian-bois" en las Guayanas, "UTA" en Perú y leishmaniasis cutánea, en general, en Costa Rica y Panamá. El uso del nombre de L. tropica se debe a la hipótesis de que proviene del botón de oriente, importado del Hemiferio Oriental, pero el hallazgo de reservorios infestados en lugares vírgenes desfavorecen este punto de vista y Pessoa da el "paso correcto" en 1961, cuando las refiere como L. braziliensis mexicana, L. braziliensis guyanensis y L. braziliensis peruviana. En 1962 Garnham dió al parásito de la úlcera del chiclero el rango específico de L. mexicana.
Ultimamente se ha venido hablando de dos grandes grupos o complejos: El L. mexicana y el L. braziliensis, conteniendo un número de especies o subespecies diferentes, basados sobre todos los criterios posibles como: morfología, biología, inmunología y epidemiología.
La taxonomía fue revisada en 1973 para incluir nuevos parásitos descubiertos asi como también nuevos criterios bioquímicos. Laison & Shaw 1972 - 1973, Brasil.
Leishmaniasis visceral: Fue en 1912 cuando esta enfermedad se observó en América, cuando Migone describe un caso en Paraguay, Mazza y Cornejo en la Argentina en 1926 y Renna en el Brasil en 1934. El agente etiológico se nombra primero como L. chagasi pero algunos han rehusado este nombre porque la clínica es similar a la del "Kala azar" del Hemiferio Oriental y, al parecer, se introdujo desde Europa, por consiguiente se debería referir como L. donovani o L. infantum. Lainson & Shaw retomaron el nombre anterior, debido a diferencias epidemiológicas, geográficas y sugieren diferentes especies responsables para la leishmaniasis visceral en América.
El parásito:
Ciclo evolutivo.- El parásito llega al vector cuando éste se alimenta de un vertebrado infestado, pasando al estómago, donde se desdiferencia, transformándose en promastigote a partir de la forma amastigote proveniente del macrófago del vertebrado. En el intestino del vector se encuentran varias formas: promastigote que se multiplica muchas veces produciendo rocetas, que originan una gran cantidad de parásitos, pasando así al tubo digestivo anterior (el proventrículo) llegando a producir una obstrucción, así el vector se encuentra imposibilitado de alimentarse y en uno de sus esfuerzos, después de una relajación de su musculatura, regurgita sangre aspirada llevando así parásitos hacia afuera inoculandolos dentro del vertebrado, una vez allí es fagocitado por los macrófagos (SRE) y en forma de amastigote se reproducen.
Lainson & Shaw (1979) agruparon a los miembros
del genero en Peripylaria y Suprapylaria basados en la posición
del desarrollo del parásito en el vector.
Los de la sección peripylaria se desarrollan en
la parte posterior del intestino antes de moverse a la
parte anterior, a esta sección pertenecen todas
las L. braziliensis y los de la sección suprapylaria
se desarrollan en el intestino medio del vector
ante de dirigirse a la parte anterior, a
esta sección pertenecen todas las otras leishmanias conocidas.
Según la clínica tenemos:
1- Exclusivamente lesiones cutáneas, botón de oriente, leishmaniasis cutánea.
2- Formas cutáneas que pueden complicarse con lesiones tegumentarias que destruyen la nariz, boca, etc., leishmaniasis tegumentaria américana.
3- Formas visceral: Estos parásitos presentan tropismos
por los órganos del SRE: bazo, hígado,
médula ósea y tejido linfoide, leishmaniasis
visceral.